
Proteine și activitate enzimatică în miere
Mierea este un amestec de cel puțin 181 de substanțe diferite [Alvarez-Suarez și colab., 2010]. Componentele mai puțin evidente ale acestui produs valoros produs de albine sunt proteinele și aminoacizii (blocuri de proteine), care, conform diferitelor surse, reprezintă de la 0,58 la chiar (în cazuri extreme) 7,86% [White, 1978].

Fot. Roman Dudzik
Fracția de proteină domină cantitativ aminoacizii - reprezintă 40 până la 80% din toți compușii de azot din miere. Prezența proteinelor face ca mierea să aibă o tensiune superficială mai mică, ceea ce determină o tendință clară de spumare și favorizează formarea de bule fine de aer [Di Girolamo și colab., 2012]. Mierea de hrișcă are cel mai mare conținut de proteine, iar cea mai mică - mierea de salcâm sau tei [Erban și colab., 2019]. Mulți ani de experimente au arătat că conținutul de proteine din miere scade cu 46,7% după 6 luni, indiferent de originea sa botanică [Brudzynski și colab., 2013].
Sursa de proteine și aminoacizi din miere sunt albinele în sine - acești compuși sunt secretați de glandele salivare și faringiene ale culegatoarelor, precum și de plante - de fapt polen, care se găsește în miere [Lewkowski și colab., 2019]. Proteinele vegetale sunt rareori găsite în profilele proteice ale mierii (cu excepția mierii de hrișcă, eucalipt, floarea soarelui și a rapiței).
Ele reprezintă de obicei o mică parte din conținutul total de proteine, deoarece pot fi digerate de proteaze de albine, filtrate (particule de polen) sau descompuse în timpul maturării mierii [Baroni și colab., 2002; Di Girolamo și colab., 2012; Erban și colab., 2019].
 [...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-Prenumeratorów
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówCompoziția proteică a mierii este, de asemenea, importantă în ceea ce privește proprietățile sale de promovare a sănătății. Acești compuși organici modelează în mod semnificativ proprietățile biocide ale mierii (distrugerea bacteriilor, virușilor, ciupercilor și protozoarelor) [Ahmed și colab., 2018; Dubiago și colab., 2018; Samarghandian și colab., 2017; Vallianou, 2014; Żegliński, 2006] și sprijinirea vindecării rănilor [așa-numitul pansamente cu miere; Irishi și colab., 2011; Weston, 2000]. Studiile anterioare privind compoziția chimică a mierii nu s-au concentrat asupra fracției proteice.
În ultimii ani, am observat o creștere a interesului pentru acest grup de compuși, iar principalul motiv este proprietățile lor semnificative pro-sănătate. Cu toate acestea, ca o consecință a acestor câteva studii, apar multe semne de întrebare cu privire la prezența proteinelor specifice (de exemplu lizozimă, transferină 1, superoxid dismutază) sau funcția lor în miere (catalază, fosfatază acidă) [Erban și colab., 2019]. Interesant este că mierea conține și proteine din veninul albinelor, inclusiv cele care provoacă reacții alergice. Erban și colab. [2019] indică faptul că mierea poate fi utilizată pentru a trata alergia la veninul de albine.
Aminoacizi
Aminoacizii joacă un rol important - sunt precursori (blocuri de construcție) care construiesc proteine, care sunt un element esențial în organism. Mierea conține 26 de aminoacizi, care constituie 0,1% până la 1% din cele menționate anterior. Printre acestea, prolina este cea dominantă, care constituie aproximativ 50-85% din toți aminoacizii. Conținutul celor 25 de aminoacizi rămași și proporțiile lor relative (așa-numitul profil de aminoacizi) depind de originea mierii (nectar sau miere).
Cu toate acestea, majoritatea mierilor de calitate se caracterizează printr-un conținut ridicat de fenilalanină, acid aspartic și acid glutamic. Mierea de cimbru este bogată în serină, tirozină și lizină. În mierea de rozmarin, tirozina este principalul aminoacid și sunt prezente și cantități mari de prolină și fenilalanină [Hermosın și colab., 2003]. Datorită faptului că principala sursă de aminoacizi din miere sunt boabele de polen, profilul de aminoacizi al mierii este caracteristic originii sale botanice [Anklam, 1998]. Originea geografică, inclusiv condițiile climatice și ale solului, poate influența, de asemenea, profilul aminoacizilor mierii [Davies, 1976].
Prolina este un aminoacid unic, deoarece originea sa în miere este de origine animală - de la albina, nu de la plantă ca majoritatea aminoacizilor. Pe lângă activitatea enzimatică, nivelul de prolină din miere este propus ca un indicator al maturității și autenticității acestui produs apicol [Von der Ohe și colab., 1991]. Pentru a fi considerată miere matură, prolina trebuie să fie mai mare de 200 mg / kg miere și cel puțin 66% din totalul aminoacizilor liberi (de obicei 80-90%).
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówEnzime ale metabolismului carbohidraților
Enzimele, care sunt, de asemenea, una dintre cele mai importante componente ale mierii în general, sunt o parte importantă a fracțiunii proteice a mierii. Sarcina lor principală este de a transforma nectarul în miere și astfel sunt responsabili pentru producerea mierii.
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówActivitatea α- și β-glucozidazei, α- și β-amilazei împreună cu nivelul de hidroximetilfurfural (HMF) sunt utilizate pentru a evalua supraîncălzirea mierii. Deși stabilitatea termică a α- și β-amilazelor este mai mare decât cea a α- și β-glucozidazelor [Berlitz și colab., 2009]. Invertazele (α- și β-glucozidaze) sunt enzime care descompun zaharoza și maltoza în zaharuri simple (glucoză și fructoză) - componenta principală a mierii. Acestea catalizează reacția principală care determină maturarea nectarului în miere.
Invertaza permite albinelor să producă o soluție foarte concentrată de zaharuri, rezistentă la fermentație, care este un aliment cu energie ridicată, care ocupă o suprafață minimă în fagure [Sánchez și colab., 2001].

Fot. Roman Dudzik
Invertazele sunt foarte sensibile la depozitarea și încălzirea mierii (de exemplu, pentru decristalizare), mai mult decât la diastaze. Încălzirea mierii sau turnarea unui lichid cu o temperatură egală sau mai mare de 50°C determină o reducere semnificativă a activității lor [Dimins și colab., 2014].
Prin urmare, în unele țări, măsurarea activității invertazei indică prospețimea mierii și condițiile corecte de depozitare. Activitatea invertazei este exprimată ca număr de invertaza (unități Hadorna), care indică cantitatea de enzimă activă conținută în miere care descompune zaharoza (exprimată în grame) în 1 oră, de enzima prezentă în 100 g de miere în condiții de testare [Hadorn și Zürcher, 1966].
Mierile proaspete și neadulterate ar trebui să aibă cel puțin 10 unități Hadorna, în timp ce mierile cu activitate enzimatică scăzută ar trebui să aibă cel puțin 4 unități [Berlitz și colab., 2009]. Mierea se caracterizează printr-o activitate ridicată a invertazelor, iar mierea de salcâm - cea mai scăzută activitate a acestui grup de enzime [Oddo și colab., 1999].
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówPractic, mierile întunecate au un număr de diastază mai mare decât mierile deschise. Cercetările efectuate au arătat că mierea de hrișcă (49), mierea de erică (43,5) și mierea de conifere (38,5) au avut cel mai mare număr de diastaze. Cel mai mic număr de diastază se găsește în mierea de salcâm (10,9), mierea de rapiță și mierea de tei (15).
Interesant este că mierile poliflore domestice se caracterizează printr-un număr mare de diastaze, adesea peste 20. Analiza Wesołowska și Dźugan [2017] arată că depozitarea mierii la o temperatură de 20 ± 2 ° C timp de 24 de luni reduce activitatea diastazei cu o medie de 31%. Cu toate acestea, cea mai importantă enzimă care, datorită activității sale, conferă mierii proprietăți antibacteriene este glucoza oxidaza. Aceasta este o enzimă pe care albinele o adaugă la nectar în timpul procesului de producere a mierii. Catalizează reacția de conversie a glucozei din nectar în acid gluconic în prezența oxigenului și a apei (Fig. 1).
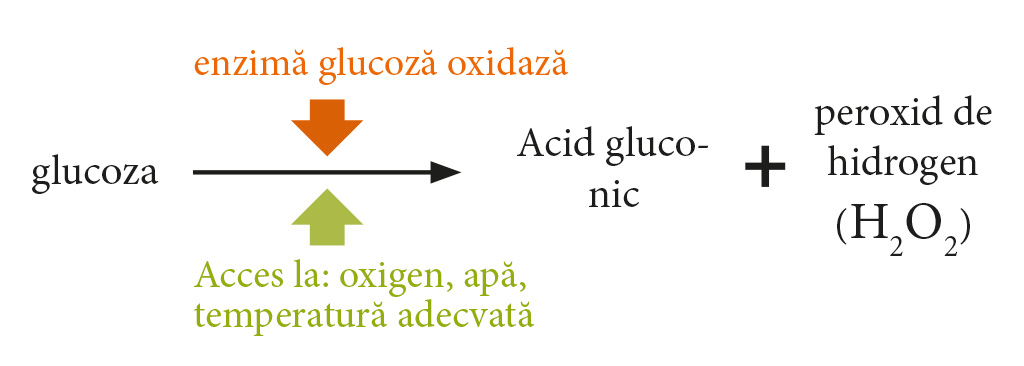
Diagrama.1. Diagrama formării peroxidului de hidrogen din glucoză. Factorii semnificativi necesari pentru efectuarea reacției sunt: prezența enzimei, accesul la oxigen, apă și temperatura adecvată a mediului de reacție. Bazat pe: Kedzia i Hołderna-Kedzia, 2017; Alb [1966].
Un produs secundar al acestei reacții este peroxidul de hidrogen (H2O2). Produsele acestei reacții (acid și H2O2) protejează mierea de contaminarea cu microorganisme în timpul maturării.
Peroxidul de hidrogen conținut în miere are, de asemenea, o altă funcție, mult mai importantă pentru potențialii consumatori - este responsabilă pentru activitatea sa biocidă împotriva bacteriilor, virușilor, ciupercilor sau protozoarelor. Studiile au arătat că moleculele de H2O2 pot pătrunde în membranele celulare ale multor agenți patogeni, unde sunt apoi descompuse de sistemele de protecție ale gazdei. Cu toate acestea, ca urmare a acestui proces, din molecula de peroxid de hidrogen distrusă se formează radicali hidroxil liberi (HO •).
Așa cum am scris în articolul precedent („Stupina24” 1/2021), radicalii liberi „iau” electroni din alți compuși chimici, inclusiv molecule de ADN, inducând mutații și deteriorând materialul genetic al gazdei. De aici și proprietățile biocide ale mierii [Ahmed și colab., 2018; Dubiago și colab., 2018; Kedzia i Hołderna-Kedzia, 2017; Rybak-Chmielewska și Szczęsna, 2008; Samarghandian și colab., 2017; Vallianou, 2014; Żegliński, 2006].
Chiar și cantități mici de peroxid de hidrogen au un efect bacteriostatic, adică limitează creșterea bacteriilor patogene: Staphylococcus aureus și Pseudomonas aeruginosa, care provoacă infecții nosocomiale și este periculoasă datorită rezistenței la numeroase antibiotice și ciuperci de drojdie. , picioare sau vagin, precum și ciuperci de mucegai Aspergillus fumigatus provocând reacții alergice sau micoză urechii [Kedzia și Hołderna-Kedzia, 2017].
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówStudii recente au arătat că această miere nu acumulează niveluri detectabile de H2O2 când este diluată cu apă, iar efectul său antibacterian se datorează prezenței unui compus organic numit metilglioxal (MGO) [Kwakman și colab., 2011]. Metilglioxal modifică enzima glucoză oxidază, schimbându-i structura și legând-o permanent, reducând în același timp producția de peroxid de hidrogen [Majtan și colab., 2014].
Al doilea grup de enzime prezente în miere include pe cele de origine vegetală - prezente în polen și nectar: catalază și fosfatază acidă [Alonso-Torre și colab., 2006; Brudzyński și colab., 2011]. Catalaza este o enzimă care se găsește în mod natural în miere. Sursa enzimei este boabele de polen și, într-o măsură mai mică, nectarul. Mai mult, activitatea catalazei de miere este parțial atribuită drojdiei și altor microorganisme [Huidobro și colab., 2005].
Nivelul de H2O2 din miere depinde de viteza de formare a acesteia - datorită activității glucozei oxidazei și a vitezei la care este descompusă de enzima catalază [Brudzyński et al., 2011)]. Funcționează prin descompunerea peroxidului de hidrogen în oxigen și apă (Fig. 2).
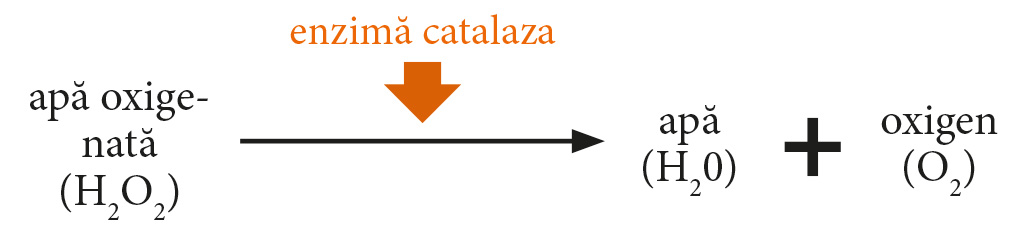
Diagrama. 2. Schema reacției catalizate de catalază.
Prin urmare, este clar că nivelul absolut de H2O2 din orice miere este determinat de nivelurile corespunzătoare de glucoză oxidază și catalază din acea miere. Cu cât este mai ridicat nivelul de glucoză oxidază, cu atât este mai mare nivelul de peroxid de hidrogen și cu cât este mai scăzut nivelul de catalază, cu atât este mai mare nivelul de peroxid de hidrogen. Este important însă, catalaza prezentă în miere nu este capabilă să descompună complet peroxidul de hidrogen format [Dustman, 1971; Weston, 2000].
Activitatea catalazei depinde de originea botanică a mierii. Astfel, cantitatea de polen colectat și sursa de origine a acestuia, precum și activitatea catalazei în polen, vor determina nivelul de peroxid de hidrogen din miere [Weston, 2000]. Mierea de salcâm, lucernă, erică și miere se caracterizează printr-o activitate catalazică mai mare comparativ cu, de exemplu, mierea de castane [Bucekova și colab., 2019; Huidobro și colab., 2015]. Activitatea catalazei de miere scade în timpul depozitării [Krauze și Krauze, 1991].
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówProteine din laptisorul de matca
O componentă proteică importantă a mierii sunt proteinele din lăptișorul de matcă [Di Girolamo și colab., 2012], secretate din glandele tinerelor lucratoare pentru a hrăni larvele și matca [Chua și colab., 2013). Acestea includ, printre altele: principalele proteine din lăptișor de matcă (MRJP) de la 1 la 8, cu cea mai mare cantitate de MRJP1, defensină-1 (rojalizină), himenoptaecină și apisimină. Aceste proteine sunt importante pentru activitatea antimicrobiană a mierii, care, împreună cu alți compuși chimici produși în timpul transformării zaharurilor (H2O2 sau acid gluconic), previn deteriorarea mierii de către microorganisme [Lewkowski și colab., 2019].
În plus, unele proteine pot stimula funcțional sistemul imunitar uman prin eliberarea unei citokine - factor de necroză tumorală din celulele imune (α-TNF) - care este implicat în mecanismele de apărare a gazdei antivirale, antibacteriene și antiparazitare [Ahmed și colab., 2018; Bilkova și colab., 2015]. Bilikova și colab. [2015] au arătat cele mai puternice proprietăți stimulatoare ale salcâmului și mierii de tei.
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówDr. Agata Nicewicz
Łukasz Nicewicz, MA
Facultatea de Științe ale Naturii
Universitatea din Silezia din Katowice
LITERATURA
Ahmed S, Sulaiman AS, Baig AA, Ibrahim M, Liagat S, Fatima S, Jabeen S, Shamim N, Othman NH, 2018. Mierea ca medicament potențial antioxidant natural: o perspectivă asupra mecanismelor sale moleculare de acțiune. Medicină oxidativă și longevitate celulară. 18: 8367846.
Alonso-Torre SR, Cavia MM, Fernández-Muiño MA, Moreno G, Huidobro JF, Sancho MT, 2006. Evoluția activității fosfatazei acide a mierilor din diferite clime. Chimia alimentelor. 97 (4): 750-755.
Anklam E, 1998. O revizuire a metodelor analitice pentru a determina originea geografică și botanică a mierii. Chimia alimentelor. 63 (4): 549-562.
Babacan S, Rand AG, 2006. Purificarea amilazei din miere. Journal of Food Science. 70 (6): c413-c418.
Baroni MV, Chiabrando GA, Costa C, Wunderlin DA, 2002. Evaluarea originii florale a mierii prin tehnici de imunoblot pe pagină SDS. J. Agric. Food Chem. 50: 1362-1367.
Berlitz HD, Grosch W, Schieberle P, 2009. Chimia alimentelor, ed. A 4-a. Springer, Leipzig
Bilkova K, Krakova TK, Yamaguchi K, Yanaguchi Y, 2015. Proteine majore din lăptișor de matcă ca markeri ai autenticității și calității mierii. ArhHigRadaToksikol. 66: 259-267.
Borawska M, Arciuch L, Puscion-Jakubiak A, Lewoc D, 2015. Conținutul de zaharuri (fructoză, glucoză, zaharoză) și prolină în diferite soiuri de miere naturală. ProblHigEpidemiol. 96 (4): 816-820.
Brudzynski K, Sjaarda C, 2015. Glicoproteine cu miere care conțin peptide antimicrobiene, jelleine ale proteinei 1 din lăptișorul de matcă major, sunt responsabile pentru activitățile litice și bactericide ale peretelui celular al mierii. Plus unu. 10 (4): e0120238.
Brudzynski K, Abubaker K, St-Martin L, Castle A, 2011. Reexaminarea rolului peroxidului de hidrogen în activitățile bacteriostatice și bactericide ale mierii. Frontiere în microbiologie. 2: 213.
Brudzynski K, Sjaarda C, Lannigan R, 2015. Glicoproteine care conțin MRJP1 izolate din miere, un medicament antibacterian nou candidat cu activitate de spectru larg împotriva izolatelor clinice rezistente la mai multe medicamente. Frontiere în microbiologie, 6.
BrudzynskiK, Lannigan R, 2012. Mecanismul acțiunii bacteriostatice a mierii împotriva MRSA și VRE implică radicali hidroxil generați din peroxidul de hidrogen al mierii. Față. Microbiol. 3: 1-8.
Brudzynski K, Sjaarda C, Maldonado-Alvarez L, 2013. O nouă privire asupra complexării proteinei-polifenolului în timpul depozitării mierii: Este acesta un eveniment aleatoriu sau organizat cu ajutorul proteinelor de tip dirigent? Plus unu. 8: e72897.
Bucekova M, Jardekova L, Juricova V, Bugarova V, Di Marco G, Gismondi A, AMajtan J, 2019. Activitatea antibacteriană a diferitelor miere de flori: descoperiri noi. Molecule. 24 (8): 1573.
Bucekova M, Sojka M, ValachovaI, Martinotti S, Ranzato E, Szep Z, Majtan V, Klaudiny J, Majtan J, 2017. Peptida antibacteriană derivată din bee, defensin-1, promovează re-epitelizarea plăgilor in vitro și in vivo. Sci.Rep. 7: 7340.
Chua LS, Lee JY, Chang GF, 2013. Extracția și determinarea proteinelor din miere prin spectrometrie de masă. Anal Bioanal Chem. 405: 3063-3074.
Chua LS, Lee YJ, Chan GF, 2013. Extracția și determinarea proteinelor din miere prin spectrometrie de masă. Chimie analitică și bioanalitică. 405 (10): 3063-3074.
Codex alimentarius, 1994. Codex Standardfor Honey, Codex Stan 121981, Rev.1 (11). FAO; Roma, Italia.
Davies AMC, 1976. Aplicarea analizei aminoacizilor la determinarea originii geografice a mierii. Journal of Food Technology. 11: 515-523.
Di Girolamo F, D'Amato A, Righetti PG, 2012. Evaluarea originii florale a mierii prin instrument proteomic. Journal of Proteomics. 75: 3699-3693.
Dimins F, Mikelsoe V, Kuka P, Jefremoves AN, 2014. Efectul diferitelor tipuri de tratament termic asupra activității invertazelor din miere. Procedee Foodbalt. 2014: 284-289.
Dubiago G, Nowak A, Klimowicz A, 2018. Proprietăți selectate ale mierii deosebit de utile în cosmetologie. PostępyFitoterapii. 1/2018: 58-64.
Dustman JH, 1967. Măsurarea peroxidului de hidrogen și a activității enzimei în mierea central-europeană. Z cercetarea albinelor. 9: 66-73.
Dustman JH, 1971. Despre activitatea catalazei în miere din costumul de erică (Ericaceae). Journal of Food Study and Research. 145: 292-295.
Dyrektywa Rady Europy 2001/110 / WEz dnia 20 grudnia 2001 r.odnosząca się do miodu. Jurnalul Oficial L 010, 12/01/2002 p. 0047-0052.
Erban T, Shcherbachenko E, Talacko P, Harant K, 2019. Compoziția unică de proteine a mierii a dezvăluit printr-o analiză proteomică cuprinzătoare: alergeni, proteine asemănătoare veninului, proprietăți antibacteriene, proteine de lăptișor de matcă, serin proteaze și inhibitorii acestora. J. Nat. Prod. 82 (5): 1217-1226.
Fagain CO, 1997. Stabilitatea proteinelor și stabilizarea funcției proteinelor. Georgetown, TX: Landes Bioscience.
Godlewska M, Świsłocka R, 2015. Proprietăți fizico-chimice și antimicrobiene ale mierii din regiunea Podlasie. Cosmos. 2 (307): 347-352.
Gonnet M., 1988. Factori naturalijeantibioticzeskije, sodierżaszcziesja în cupru. [În:] Produsele bărcii de pescuit - scârțâituri, sănătate, frumusețe. Izd. Apimondii, București 33-7.
Grecka K, Kus PM, Worobo RW, Szweda P, 2018. Studiul potențialului anti-stafilococic al mierilor produse în nordul Poloniei. Molecule. 23: 260.
Hadorn H, Zürcher K, 1966. EineverbessertepolarimetrischeMethodezurSaccharasezahlbestimmungimHonig. Dt. Lebensm. Rdsch. 62: 195-201.
Hermosın I, Chicón RM, Cabezudo DM., 2003. Compoziție de aminoacizi liberi și originea botanică a mierii. Food MP, Muniategui S, Sancho MT, 2005. Metodă precisă pentru măsurarea activității catalazei în miere. J. AOAC Int. 88: 800-804.
Irish J, Blair S, Carter DA, 2011. Activitatea antibacteriană a mierii derivate din flora australiană. Plus unu. 6 (3): e18229.
Janiszewska K, Aniołowska M, Howis M, Nowakowski P, 2011. Compoziția aminoacizilor liberi ca marker al sursei de origine a mierii. Industria chimica. 90 (5): 806-808.
Kędzie B, Hołderna-Kezja E, 2017. Opinii contemporane asupra mecanismului acțiunii antimicrobiene a mierii. Post Phytoter. 18 (4): 290-297.
Kowalski S, Lukasiewicz M, Bednarz S, Panuś M, 2012. Numărul diastazei se modifică în timpul procesării termice și cu microunde a mierii. Ceh J. Food Sci. 30 (1): 21-26.
Krauze A, KrauzeJ, 1991. Modificări ale compoziției chimice a mierilor stocate. Acta Aliment. Pol. 2 (119): 126 –140.
Kretavicius J, Kurtinaitiene B, Racys J, Čeksteryte V, 2010. Inactivarea glucozei oxidazei în timpul descristalizării mierii prin tratamentul termic. Zemdirbyste. 97 (4): 115-122.
Kwakman PH, TeVelde AA, De Boer L, Vandenbroucke-Grauls CM, Zaat SA, 2011. Două mieri medicinale majore au mecanisme diferite de activitate bactericidă. Plus unu. 6: e17709.
Kwakman PHS, Boer L, Ruyter-Spira CP, Creemers-Molenaar T, Helsper JPFG, Vandenbroucke-Grauls CMJE, Zaat SAJ, Velde AA, 2011. Miere medicală îmbogățită cu peptide antimicrobiene a îmbunătățit activitatea împotriva agenților patogeni rezistenți la antibiotice. Eur JClinMicrobiol Infect Dis. 30: 251-7.
Lee DC, Lee SY, Cha SH, Choi YS, Rhee HI, 1998. Discriminarea mierii de albine native și a mierii de albine străine prin SDS-PAGE. Coreeană J Food Sci. 30: 1-5.
Lewkowski O, Muresan CI, Dobrizsch D, Fuszard M, Erler S, 2019. Efectul dietei asupra compoziției și stabilității proteinelor secretate de albine în miere. Insecte. 10: 282.
Majewska E, Drużyńska B, Derewiaka D, Cicierska M, Wołosiak R, 2015. Calități fizico-chimice ale mierilor de nectar selectate. Bromat. Chem. Toxicol. XLVIII, 3: 440-444.
Majtan J, Bohova J, Prochazka E, Kaludiny J, 2014. Metilglioxal poate afecta acumularea de peroxid de hidrogen în miere de manuka prin inhibarea glucozei oxidazei. J Med Food. 17 (2): 290-292.
Majtan J, Majtan V, 2010. Este mierea de manuka cel mai bun tip de miere pentru îngrijirea rănilor? J Hosp Infect. 73: 305–306.
Oddo LP, Piazza MG, Pulcini P, 1999. Activitatea inversării în miere. Apidologie. 30: 57-65.
PN-88 / A-77626. Polska Norma - Miód pszczeli, 1988. Polski Komitet Normalizacji, Miar i Jakości. Warszawa: WydawnictwaNormalizacyjne Alfa.
Pontoh J, NH scăzut, 2002. Purificarea și caracterizarea beta-glucozidazei de la albinele de miere (Apis mellifera). Insecte Biochem. Mol. Biol. 32: 679-690.
Ordonanța ministrului agriculturii și dezvoltării rurale din 14 ianuarie 2009 privind metodele de analiză legate de evaluarea mierii. Journal of Laws 2009 Nr. 17 articol 94.
Rybak-Chmielewska H, Szczęsna T, 2008. Produse apicole. [În:] Wilde J., Prabucki J. (eds.), Hodowla honey bees. PWRi, Poznanń, pp. 323-349.
Samarghandian S, Farkhondeh T, Samini F, 2017. Miere și sănătate: o revizuire a cercetărilor clinice recente. Cercetare în farmacognozie. 9 (2): 121-127.
Sánchez MP, Huidobro JF, Mato I, Muniategui S, Sancho MT, 2001. Evoluția activității inversazelor în miere în doi ani. Jurnalul de chimie agricolă și alimentară. 49 (1): 416-422.
Stagos D, Soulitsiotis N, Tsadila C, Papaeconomou S, Arvanitis C, Ntontos A, Karkanta F, Adamou-Androulaki S, Petrotos K, Spandidos DA, 2018. Activitate antibacteriană și antioxidantă a diferitelor tipuri de miere derivate din Muntele Olimp din Grecia. Int. J. Mol. Med. 42: 726-734.
Vallianou N, 2014. Mierea și proprietățile sale antiinflamatorii, anti-bacteriene și anti-oxidante. Medicina generala. 2: 2.
Von der Ohe W, Dustmann JH, Von der Ohe K, 1991. ProlinalsKriterium der Reife des Honigs. Deutsche Lebensmittel-Rundschau. 87 (12): 383-386.
Wesołowska M, Dżugan M, 2017. Activitatea și stabilitatea termică a diastazei care apar în mierile varietale din Podkarpacie. Alimente. Ştiinţă. Tehnologie. Calitate. 24 (4): 103-112.
WestonRJ, 2000. Contribuția catalazei și a altor produse naturale la activitatea antibacteriană a mierii: o revizuire. Food Chem. 71: 235-239.
White JW, 1957. Compoziția mierii. Bee World. 38: 57-66.
White JW, 1966. Inhibină și glucoză oxidază în miere - o recenzie. Am Bee J. 106: 214-6.
White JW, 1978. Dragă. Adv. Alimente Res. 24: 287-374.
Câștigat SR, Lee DC, Ko SH, Kim JW, Rhee HI, 2008. Caracterizarea proteinei majore a mierii și aplicarea acesteia la detectarea adulterării. Food Res Int, 41: 952-956.
Żegliński J, 2006. Stabilizarea peroxidului de hidrogen în xerogelosilicat - studiul interacțiunii componentelor compozite și a caracteristicilor sale. Teză de doctorat. Departamentul de Chimie Anorganică, Universitatea de Medicină din Gdańsk. Str.: 63.
Zhang YZ, Chen YF, Wu YQ, Si JJ, Zhang CP, Zheng HQ, Hu FL, 2019. Discriminarea originii entomologice a mierii în funcție de secrețiile albinei (Apiscerana sau Apis mellifera). Int. 116: 362-369.
 AbonamentГодовая
AbonamentГодовая
подпискаZamów prenumeratę czasopisma "Pasieka"
