


Rośliny inwazyjne – wsparcie czy zagrożenie dla owadów zapylających?
Inwazyjne gatunki obce wymieniane są obecnie wśród czynników stanowiących największe zagrożenie dla różnorodności biologicznej [Luque i in. 2014].
Destabilizacja biocenoz, będąca skutkiem działalności człowieka, m.in. intensyfikacji produkcji rolniczej, zmniejszaniu powierzchni obszarów nieprodukcyjnych oraz urbanizacja, sprzyja pojawianiu się obcych gatunków roślin, których rozprzestrzenianie się jest trudne do opanowania [Paszkiewicz-Jasińska i Żyszkowska 2011].
Rośliny te nie tylko powodują zmiany składu i stosunków ilościowych gatunków w zbiorowiskach, ale wywołują również ogromne straty w gospodarce oraz zagrażają zdrowiu człowieka [Heink i in. 2018].

Fot. Jacek Jachuła
Zmniejszenie populacji owadów zapylających, zarówno pszczoły miodnej, jak i owadów dziko żyjących, jest problemem globalnym. W strefie klimatu umiarkowanego owady zapylają około 80% gatunków roślin. Obecnie, w wielu regionach świata stwierdza się niedobory zapylaczy.
Zjawisko to wywołuje straty w produkcji rolniczej oraz ograniczenie bioróżnorodności ze względu na obniżenie sukcesu reprodukcyjnego gatunków uprawnych i dziko rosnących [Majewski 2011].
 [...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-Prenumeratorów
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówOddziaływanie na florę rodzimą
Wypieranie flory autochtonicznej odbywa się zazwyczaj w wyniku konkurencji o zasoby środowiska, głównie światło i wodę. Pędy roślin inwazyjnych mogą osiągać znaczną wysokość i powodować zacienianie gatunków o niższym pokroju.
Wysoka skuteczność rozmnażania wegetatywnego, szybki przyrost biomasy związany ze zwiększonym poborem wody i zdolność do regeneracji uszkodzonych organów skutkuje ograniczeniem przestrzeni życiowej gatunków rodzimych i prowadzi do monotypizacji gatunkowej siedlisk [Tokarska- Guzik i in. 2012].
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówTab. 1. Najważniejsze cechy roślin obcych decydujące o ich inwazyjności
|
Cecha |
Przykłady gatunków |
|
wysoka płodność, trwałość nasion, przedłużona zdolność kiełkowania |
klon jesionolistny, niecierpek drobnokwiatowy |
|
budowa propagul umożliwiająca wykorzystanie zewnętrznych czynników transportowych (niski ciężar, niezatapialność, atrakcyjność dla zwierząt zjadających zewnętrzną część owoców) |
klon jesionolistny, niecierpek drobnokwiatowy, czeremcha amerykańska, wiele gatunków z rodziny astrowatych (Asteraceae) |
|
plastyczność morfologiczna, wczesne przystępowanie do reprodukcji, znaczna alokacja biomasy w część podziemną |
klon jesionolistny, rdestowiec japoński |
|
dwupienność, poligamia, samopylność fakultatywna, przyspieszona dezintegracja osobnika macierzystego na osobniki potomne |
klon jesionolistny |
|
zdolność do konkurowania z innymi gatunkami (wzrostem, tempem odtwarzania uszkodzonych organów) |
czeremcha amerykańska, robinia akacjowa, rdestowiec sachaliński, barszcz Sosnowskiego |
|
zmienność fenotypowa, zdolność do tworzenia mutantów, poliploidów, mieszańców z pokrewnymi gatunkami w nowej ojczyźnie |
gatunki wiesiołków, przymiotna, szczaw omszony |
Źródło: [Kołaczkowska 2008]
Wpływ na sieci zapyleń
Wpływ roślin inwazyjnych na interakcje rodzimych gatunków roślin z owadami zapylającymi stał się przedmiotem intensywnych badań [Bjerknes i in. 2007]. Oddziaływanie to może być pozytywne (przyciąganie zapylaczy do zbiorowisk), negatywne (zmniejszenie frekwencji wizyt owadów na kwiatach roślin rodzimych) lub neutralne.
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówTab. 2. Zróżnicowanie wpływu Carpobrotus acinaciformis na florę rodzimą w zależności od lokalizacji
|
Gatunek inwazyjny |
Gatunek rodzimy |
Lokalizacja |
Rodzaj oddziaływania |
|
Carpobrotus acinaciformis |
Lotus cytisoides |
Baleary |
konkurencja o zapylacze |
|
Cistus salviifolius, |
przyciąganie owadów zapylających |
||
|
Cistus monspeliensis |
brak wpływu na liczbę wizyt owadów |
||
|
Dithyrea maritima, |
Kalifornia, USA |
brak wpływu na liczbę wizyt owadów |
|
|
Helichrysum stoechas |
Majorka |
negatywny wpływ na wiązanie nasion |
Źródło: [Stout i Morales 2009; zmienione].
Efekt, jaki wywiera wkroczenie gatunku obcego do flory rodzimej na sieć zapyleń, może także wyraźnie różnić się pomiędzy kolejnymi latami obserwacji. Obecność wilczomlecza lancetowatego (Euphorbia esula L.) w zbiorowisku z dzwonkiem okrągłolistnym (Campanula rotundifolia L.) w USA początkowo powodowała zwiększenie frekwencji odwiedzin owadów w kwiatach C. rotundifolia, lecz w kolejnym roku wywołała znaczny jej spadek [Larson i in. 2006].
Rośliny inwazyjne zagrażają w szczególności zapylaczom-specjalistom, które ze względu na specyficzne wymagania pokarmowe oraz cechy morfologiczne kwiatów roślin inwazyjnych, nie są w stanie korzystać z pożytku oferowanego przez gatunki obce.
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówWielkość pożytku oferowanego przez gatunki inwazyjne
Owadopylne rośliny inwazyjne, ze względu na zazwyczaj obfite i często długie kwitnienie oraz znaczną ilość oferowanego pożytku, są atrakcyjne dla zapylaczy [Ghazoul 2002]. Wydajność pyłkową czterech najbardziej rozpowszechnionych w Polsce gatunków inwazyjnych roślin zielnych: rukiewnika wschodniego (Bunias orientalis L.), słonecznika bulwiastego (Helianthus tuberosus L.), nawłoci kanadyjskiej (Solidago canadensis L.) i nawłoci olbrzymiej (Solidago gigantea Ait.) w warunkach Lublina określiła Denisow [2011].
Najwięcej pyłku, tj. 69,92 kg/ha dostarczał rukiewnik wschodni. Nieznacznie niższą wydajność pyłkową obliczono dla nawłoci olbrzymiej i słonecznika bulwiastego (ryc. 1.).
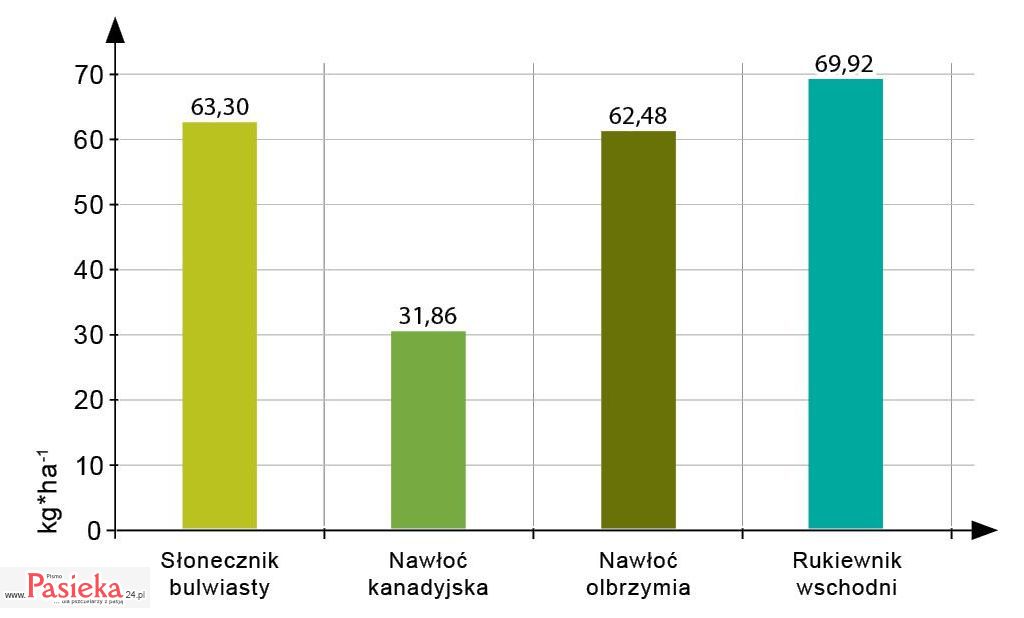
Ryc. 1. Wydajność pyłkowa wybranych roślin inwazyjnych w warunkach Lublina (średnia z lat 2006-2010)
Źródło: [Denisow 2011; zmienione]
Dodatkowo oba gatunki nawłoci i słonecznik bulwiasty kwitły aż do jesieni, wypełniając lukę w taśmie pokarmowej owadów zapylających (ryc. 2.).
Ryc. 2. Fenologia kwitnienia wybranych roślin inwazyjnych w warunkach Lublina (średnia z lat 2006-2010)
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-Prenumeratorów10% - początek kwitnienia (10% rozwiniętych pąków)
70% - pełnia kwitnienia (70% rozwiniętych pąków)
90% - końcowe stadium kwitnienia (90% przekwitających kwiatów)
Źródło: [Denisow 2011; zmienione].
Nawłoć pospolita oraz nawłoć olbrzymia są ponadto opisywane jako rośliny o wysokiej wydajności miodowej, sięgającej 700–800 kg/ha. Gatunkami inwazyjnymi dostarczającymi sporej ilości pożytku owadom zapylającym są również niecierpek gruczołowaty, trojeść amerykańska, czy naparstnica purpurowa (tab. 3.) [Kołtowski 2006].
Tab. 3. Wydajność pyłkowa i miodowa wybranych roślin inwazyjnych w Polsce
|
Gatunek rośliny |
Wydajność pyłkowa |
Wydajność miodowa |
|
Ailanthus altissima (Mill.) Swingle |
20 |
40 |
|
Asclepias syriaca L. |
brak danych |
600 |
|
Cotoneaster divaricatus |
5–7 |
190 |
|
Digitalis purpurea L. naparstnica purpurowa |
10–30 |
200 |
|
Heracleum sosnowskyi L. barszcz Sosnowskiego |
brak danych |
300 |
|
Impatiens glandulifera Royle niecierpek gruczołowaty |
400–600 |
700 |
|
Padus serotina (Ehrh.) Borkh. czeremcha amerykańska |
20 |
15 |
|
Reynoutria japonica Houtt. rdestowiec ostrokończysty |
brak dokładnych danych |
250 |
|
Rhus typhina L. sumak octowiec |
30 |
2-4 |
|
Robinia pseudacacia L. robinia akacjowa |
15 |
65 |
Źródło: [Kołtowski 2006, Tokarska-Guzik i in. 2012; zmienione].
Jakość nagrody kwiatowej
Skład oferowanego przez gatunki inwazyjne pożytku, głównie zawartość aminokwasów i białka w pyłku oraz cukrów i aminokwasów w nektarze ma bezpośredni wpływ na intensywność oraz wzorce oblotu przez rodzime gatunki zapylaczy.
Inwazyjne gatunki roślin dostarczające pyłek i/lub nektar bogaty w niezbędne poszczególnym grupom zapylaczy składniki odżywcze, stanowią uzupełniające źródło pokarmu, szczególnie w silnie przekształconym krajobrazie rolniczym [Stout i Morales 2009].
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-Prenumeratorów

Fot. Karolina Tymoszuk
Pożytek dostarczany przez niektóre rośliny inwazyjne zawiera szkodliwe dla owadów zapylających składniki. W nektarze produkowanym w kwiatach inwazyjnego dla flory Wielkiej Brytanii różanecznika pontyjskiego (Rhododendron ponticum L.) stwierdza się obecność grajanotoksyny (rodotoksyny, andromedotoksyny), substancji toksycznej dla pszczoły miodnej (Apis mellifera L.) [Koca i Koca 2007].
Tiedeken i in. [2014] wykazali, że spożycie nektaru R. ponticum przez A. mellifera spowodowało śmierć 80% obserwowanych osobników już po 6 godzinach. W przypadku pszczół samotnic (rodzaj Andrena Fabr.), odnotowywano apatyczne zachowanie i ogólne osłabienie. Co ciekawe, trzmiele (np. trzmiel gajowy Bombus lucorum L. czy trzmiel ziemny Bombus terrestris L.) wykazują odporność na działanie tej toksyny (brak efektu letalnego, subletalnego czy wpływu na zachowanie), zatem mogą korzystać z pożytku dostarczanego przez różanecznik pontyjski.
Wśród roślin inwazyjnych, których nektar zawiera toksyczne dla pszczół metabolity wtórne, wymienia się także np. obce dla flory Stanów Zjednoczonych psiankę czarną (Solanum nigrum L.), czy Corynocarpus laevigata J. R. & G. Forst [Adler 2000].
Oddziaływanie na populacje owadów
Pojawienie się inwazyjnych gatunków roślin wpływa na zapylacze nie tylko na poziomie osobniczym. Istotne jest określenie ich bezpośredniego oddziaływania na populacje tych owadów. Badań dotyczących tego zagadnienia jest niestety niewiele.
Wpływ gatunku inwazyjnego – wilczomlecza lancetowatego (Euphorbia esula L.) – na sukces reprodukcyjny pszczoły z grupy samotnic – murarki Osmia lignaria Say – badali w USA Paladini i Maron [2014].
[...] - część treści ukryta, w całości dostępna tylko dla zalogowanych e-PrenumeratorówPodsumowanie
Przemiany zachodzące w środowisku naturalnym, będące skutkiem antropopresji, spowodowały wzrost zagrożenia dla równowagi biologicznej ze strony inwazyjnych gatunków roślin. Rośliny inwazyjne w sposób pośredni (np. kształtowanie sieci zapyleń) lub bezpośredni (zmiana diety) wpływają na owady zapylające.
W wielu badaniach opisuje się zarówno pozytywny, jak i negatywny wpływ roślin inwazyjnych na rodzimą entomofaunę zapylaczy. Rodzaj oddziaływania zależy od wielu czynników, np. składu gatunkowego zbiorowiska roślinnego, liczebności populacji poszczególnych gatunków flory, gatunków owadów odwiedzających kwiaty.
Zgromadzone dane w literaturze odnoszą się jednak zazwyczaj do krótkotrwałych efektów pojawienia się gatunków inwazyjnych. W wielu przypadkach nie sposób również ocenić ich oddziaływanie na entomofaunę jako jednoznacznie pozytywne lub negatywne. Potrzeba zatem dalszych badań, które pomogą określić wpływ inwazyjnych gatunków roślin na owady zapylające.
Jacek Jachuła
Uniwersytet Przyrodniczy w Lublinie
Katedra Botaniki i Fizjologii Roślin
Studenckie Koło Naukowe Biologów
Karolina Tymoszuk
Uniwersytet Przyrodniczy w Lublinie
Katedra Botaniki i Fizjologii Roślin
Studenckie Koło Naukowe Biologów
Źródło artykułu: E.M. Szymański (red.) Ochrona pszczół i pszczelarstwo w badaniach młodych naukowców, Wydawca Górnołużyckie Stowarzyszenie Pszczelarzy w Zgorzelcu, Zgorzelec 2016, s. 92-102.
Artykuł uaktualniono i uzupełniono.
Bibliografia
Adler L.S., The ecological significance of toxic nectar, w: “Oikos” nr 91(3), 2000, s. 409-420,
Biesmeijer J.C., Roberts S.P.M., Reemer M., Ohlemueller R., Edwards M., Peeters T., Schaffers A.P., Potts S.G., Kleukers R., Thomas C.D., Settele J., Kunin W.E., Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands, w: “Science” nr 313, 2006, s. 351-354,
Bjerknes A.L., Totland O., Hegland S.J., Nielsen A., Do alien plant invasions really affect pollination success in native plant species, w: “Biol. Conserv.” nr 138, 2007, s. 1-12,
Chittka L., Schurkens S., Successful invasion of a floral market, w: “Nature” nr 411, 2001, s. 653,
Denisow B., Pollen production of selected ruderal plants species in the Lublin area, Rozprawa habilitacyjna, Uniwersytet Przyrodniczy w Lublinie, 2011,
Dostál P., Müllerová J., Pyšek P., Pergl J., Klinerová T., The impact of an invasive plant changes over time, w: ”Ecology Letters” nr 16(10), 2013, s. 1277-1284,
Ghazoul J., Flowers at the front line of invasion?, w: “Ecol. Entomol.” nr 27, 2002, s. 638-640,
Gniazdowska A., Oddziaływania allelopatyczne – ,,nowa broń” roślin inwazyjnych, w: „Kosmos” nr 54.2-3, 2005, s. 221-226,
Goulson D., Nicholls E., Botias C., Rotheray E. L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers, w: “Science” nr 347 (6229), 2015,
Heink U., Van Herzele A., Bela G., Kalóczkai Á., Jax K. Different arguments, same conclusions: how is action against invasive alien species justified in the context of European policy?, w: ”Biodiversity and Conservation” nr 27(7), 2018, s. 1659-1677,
Iacarella J. C., Mankiewicz P. S., Ricciardi A., Negative competitive effects of invasive plants change with time since invasion, w: ”Ecosphere” nr 6(7), 2015, s. 1-14,
Jakobsson A., Padrón. B., Does the invasive Lupinus polyphyllus increase pollinator visitation to a native herb through effects on pollinator population sizes?, w: “Oecologia” nr 174 (1), 2014, s. 217-226,
Koca I., Koca A.F., Poisoning by mad honey: A brief review, w: “Food Chemistry and Toxicology” nr 45, 2009, s. 1315-1318,
Kołaczkowska E., Inwazje obcych gatunków roślin – problem naukowy i praktyczny, w: „Przegląd Geograficzny” nr 54, 2008, s. 55-73,Kostuch J., Kostuch R., Rośliny inwazyjne, w: „Wieś i doradztwo” nr 1-2 (73-74), 2013, s. 21-25,
Larson D.L., Royer R.A., Royer M.R., Insect visitation and pollen deposition in an invaded prairie plant community, w: “Biol. Conserv.” nr 130, 2006, s. 148-159,
Larsson M., To Bee or Not to Be – Critical Floral Resources of Wild-Bees, Rozprawa doktorska, Uppsala University, 2006,
Łopucki R., Mróz I., Abiotyczne zmiany środowiska wywołane przez rudbekię nagą Rudbeckia laciniata (L.) – inwazyjny gatunek rośliny z rodziny astrowatych Asteraceae, w: „Studia i Materiały Centrum Edukacji Przyrodniczo-Leśnej” nr 14.4 (33), 2012, s. 241-249,
Luque G.M., Bellard C., Bertelsmeier C., Bonnaud E., Genovesi P., Simberlof, D., Courchamp F., The 100th of the world’s worst invasive alien species, w: “Biological Invasions” nr 16(5), 2014, s. 981-985,
Majewski J., Wartość zapylania upraw w województwie mazowieckim, próba szacunku, w: „Zeszyty Naukowe SGGW w Warszawie – Problemy Rolnictwa Światowego” nr 11(26), 2011, s. 112-120,
Moroń D., Skórka P., Lenda M., Kajzer-Bonk J., Mielczarek Ł., Rożej-Pabijan E., Wantuch, M., Linear and non-linear effects of goldenrod invasions on native pollinator and plant populations, w: ”Biological Invasions” nr 21(3), 2019, s. 947-960,
Nienhuis C.M., Dietzsch A.C., Stout J.C., The impacts of an invasive alien plant and its removal on native bees, w: “Apidologie” nr 40(4), 2009,
Palladini J.D., Maron J.L., Reproduction and survival of a solitary bee along native and exotic floral resource gradients, w: “Oecologia” nr 176(3), 2014, s. 789-798,
Paszkiewicz-Jasińska A., Żyszkowska M., Charakterystyka i ocena stopnia synantropizacji zbiorowisk łąkowych Gór Sowich (Sudety Środkowe), w: „Acta Botanica Silesiaca” nr 7, 2011, s. 37-47,
Sekutowski T.R., Bortniak M., Domaradzki K., Assessment of allelopathic potential of invasive plants–goldenrod (Solidago gigantea) on buckwheat (Fagopyrum sagittatum) and sunflower (Helianthus annuus), w: “Journal of Research and Applications in Agricultural Engineering” nr 57.4, 2012, s. 86-91,
Stout J.C., Morales C.L., Ecological impacts of invasive alien species on bees, w: “Apidologie” nr 40(3), 2009, s. 388-409,
Stout J.C., Tiedeken E.J., Direct interactions between invasive plants and native pollinators: evidence, impacts and approaches w: ”Functional Ecology” nr 31(1), 2017, s. 38-46,
Tokarska-Guzik B., Dajdok Z., Zając M., Zając A., Urbisz A., Danielewicz W., Hołdyński C., Rośliny obcego pochodzenia w Polsce ze szczególnym uwzględnieniem gatunków inwazyjnych, Generalna Dyrekcja Ochrony Środowiska, Warszawa, 2012, s. 73-79.
